亂僩僺僢僋僗亃
僶僀僆儅僗摐壔偵偍偗傞摐幙寢崌儌僕儏乕儖 (CBM) 偺摿惈偲墳梡
姟揷廋堦
嶰廳戝堾丒抧堟僀僲儀乕僔儑儞妛
侾丏偼偠傔偵
怉暔嵶朎暻偵娷傑傟傞峔憿懡摐椶偼丄偙偺抧媴忋偱嵟傕懡偔惗嶻偝傟傞桳婡暔幙偱偁傝丄偙傟傪嵞惗壜擻側僄僱儖僊乕尮丄偁傞偄偼丄帒尮偲偟偰棙梡偡傞偙偲偼丄恖椶偺挿擭偺柌偱偁傞丅帺慠奅偱偼丄偙傟傜偺峔憿懡摐椶偑旝惗暔偵傛傝暘夝偝傟偰偍傝丄惗暔寳偺扽慺僼儘乕偵偍偄偰旝惗暔偼廳梫側栶妱傪壥偨偟偰偄傞丅偙傟傜偺旝惗暔偑丄偳偺傛偆偵怉暔嵶朎暻傪峺慺暘夝偟偰偄傞偐偲偄偆儊僇僯僘儉傪夝柧偡傞偙偲偼丄偙傟傜偺棙梡傪幚尰偡傞偨傔偵廳梫偱偁傝丄幚嵺偵丄偙傟傑偱偵朿戝側尋媶偑峴傢傟偰偒偨丅杮僩僺僢僋偱偼丄怉暔嵶朎暻懡摐椶傪暘夝偡傞峺慺偑傕偮摿挜偺傂偲偮偱偁傞丄摐幙寢崌儌僕儏乕儖 (CBM) 偵偮偄偰徯夘偟偨偄丅偨偩偟丄尰嵼CBM偼丄摐傪擣幆偟偰寢崌偡傞僞儞僷僋幙僪儊僀儞偲偟偰峀偔掕媊偝傟偰偍傝丄摦暔偵偍偗傞摐嵔擣幆偵娭偡傞傕偺傕娷傑傟偰偄傞丅杮僩僺僢僋偱偼丄摿偵怉暔嵶朎暻懡摐椶偵寢崌偡傞傕偺偵偮偄偰徯夘偡傞丅
俀丏摐幙寢崌儌僕儏乕儖 (CBM)
僙儖儘乕僗傪壛悈暘夝偡傞僙儖儔乕僛偑懠偺峺慺偲堎側傞偺偼丄婎幙偱偁傞僙儖儘乕僗偑悈偵晄梟惈偺婎幙偱偁傞偙偲偱偁傞丅偟偨偑偭偰丄僙儖儔乕僛偼丄偙傟傪壛悈暘夝偡傞偨傔偵丄偦偺婎幙昞柺偵寢崌偟側偗傟偽側傜側偄丅Trichoderma reesei桼棃偺僙儖儔乕僛 (僙儘價僆僸僪儘儔乕僛Cel7A) 傪僷僷僀儞偱壛悈暘夝偡傞偙偲偱丄峺慺傪怗攠嶌梡偺偁傞僪儊僀儞偲丄婎幙寢崌僪儊僀儞偵暘偗傞偙偲偑偱偒偨丅怗攠僪儊僀儞偼丄婎幙寢崌僪儊僀儞傪庢傝彍偔偲丄壜梟惈偺婎幙偵懳偡傞妶惈偵偼曄壔偑側偄傕偺偺丄寢徎惈僙儖儘乕僗偵懳偡傞峺慺妶惈偑幐傢傟傞偙偲偑帵偝傟偨1)丅偙偺傛偆偵丄僙儖儔乕僛偺堦晹暘偱偁傞僙儖儘乕僗寢崌僪儊僀儞偼丄偦偺傾儈僲巁攝楍偐傜丄偄偔偮偐偺僌儖乕僾偵暘偗傜傟偰偒偨2)丅尰嵼偱偼僙儖儘乕僗寢崌僪儊僀儞偼丄峀偔摐幙傪昗揑偲偟偰寢崌偡傞僪儊僀儞偲偟偰丄乽摐幙寢崌儌僕儏乕儖 (Carbohydrate-binding module) (CBM) 乿偲柦柤偝傟丄CAZy僨乕僞儀乕僗 (http://www.cazy.or/) 忋偵婰嵹偝傟偰偄傞丅栚壓偺偲偙傠CBM僼傽儈儕乕偼丄64傑偱掕媊偝傟偰偍傝丄峏怴偑懕偄偰偄傞3)丅
偙偺僨乕僞儀乕僗偱偼丄傾儈僲巁攝楍偺傒偑峫椂偝傟僼傽儈儕乕偑掕媊偝傟偰偄傞偺偱丄摨偠僼傽儈儕乕偵懏偡傞CBM偵偍偄偰傕丄寢崌婎幙偑堎側偭偰偄傞応崌傕偁傞丅曋媂忋丄CBM偺僼傽儈儕乕偺応崌丄CBM傪棯崋偲偟丄偦偙偵僼傽儈儕乕偺斣崋傪婰弎偡傞丅CBM1偼丄僼傽儈儕乕1偵懏偡傞摐幙寢崌儌僕儏乕儖傪帵偟丄CBM30偼丄僼傽儈儕乕30偵懏偡傞傕偺傪帵偟偰偄傞丅僙儖儔乕僛 (僄儞僪僌儖僇僫乕僛丄僄僉僜僌儖僇僫乕僛偵懏偟丄EC3.2.1.4丄3.2.1.91丄3.2.1.176偺妶惈傪帩偮峺慺) 偑傕偭偰偄傞CBM僼傽儈儕乕偼丄1丄2丄3丄4丄5丄6丄8丄10丄11 丄17丄18丄28丄30丄32丄35丄37丄44丄46丄49丄63偲峀斖埻偵懏偟偰偄傞丅CBM偺僐乕僪偝傟偰偄傞埵抲偼丄N枛抂懁丄C枛抂懁丄偁傞偄偼暋悢偺怗攠僪儊僀儞偺娫偵偁傞応崌傕偁傝丄峺慺偵傛偭偰堎側傞丅傑偨丄CBM偑僞儞僨儉偵暋悢暲傫偱偄傞応崌傕偁傞丅傑偨丄摿掕偺摐幙壛悈暘夝峺慺僼傽儈儕乕 (GH) 偲偺慻傒崌偣偱懚嵼偟偰偄傞応崌傕偁傞丅偨偲偊偽丄GH10偵懏偡傞僉僔儔僫乕僛偲CBM9偲偺慻傒崌偣偑抦傜傟偰偄傞丅
昞1偵帵偡傛偆偵丄CBM1傪傕偮峺慺偼懡條偱偁傞丅偟偐偟丄偄偢傟傕怉暔嵶朎暻懡摐椶偵娭楢偡傞峺慺偱偁傝丄僙儖儘乕僗偵寢崌偡傞偲偄偆摿惈偑丄偙傟傜偺峺慺傪怉暔嵶朎暻偵廤傔偰偄傞丅峺慺杮棃偺暘夝婎幙偵寢崌偟側偔偰傕僙儖儘乕僗偵寢崌偡傞偙偲偱丄嵶朎暻懡摐椶偺暘夝偵偍偄偰偼廫暘偵岠壥傪敪婗偡傞偲峫偊傜傟傞4)丅CBM1偼丄恀妀惗暔偵摿桳偱丄堦曽丄CBM4丄10丄11 丄12丄17側偳偼丄尨妀惗暔偺傒偵尷掕偝傟偰偄傞丅傑偨CBM37偼丄懏摿堎揑偱丄偙傟傑偱偺偲偙傠丄Rumiococcus懏偵偟偐尒偮偐偭偰偄側偄丅CBM43傗CBM49偼丄怉暔摿堎揑偱偁傞丅
昞1丂摐幙寢崌儌僕儏乕儖 (CBM1) 傪帩偭偰偄傞峺慺

俁丏CBM偺寢崌條幃偲僞儞僷僋幙峔憿
摐幙傊偺寢崌條幃偵傛傝丄CBM傪僞僀僾偵傢偗傞偙偲偑丄Boraston傜偵傛傝採埬偝傟丄偙傟偑峀偔庴偗擖傟傜傟偰偄傞5)丅僞僀僾A偺CBM偼丄僼儔僢僩側寢崌柺傪傕偪丄寢崌柺偵偼丄僩儕僾僩僼傽儞傗僠儘僔儞偲偄偭偨朏崄娐傪傕偮傾儈僲巁偑暲傫偱偄傞丅偙傟傜偺傾儈僲巁偺慳悈揑側憡屳嶌梡偵傛傝丄僙儖儘乕僗偺寢徎昞柺偵寢崌偡傞丅僼傽儈儕乕偲偟偰偼丄1丄2丄3丄5丄10側偳偑抦傜傟偰偄傞丅僞僀僾B偼丄僞儞僷僋幙昞柺偵僋儗僼僩偲屇偽傟傞婎幙傪寢崌偡傞峚偑偁傞丅偟偨偑偭偰丄峚偵擖傞傛偆側梀棧偟偨摐嵔偑寢崌偵晄壜寚偱偁傞丅僋儗僼僩撪偵偼丄朏崄懓傾儈僲巁巆婎傗嬌惈傪傕偮傾儈僲巁偑攝抲偝傟偰偍傝丄峺慺偺僒僽僒僀僩偺傛偆側峔憿傪傕偭偰偄傞丅傎偲傫偳偺CBM偼丄偙偺僞僀僾B偵懏偟偰偄傞丅僞僀僾C偼丄抁偄僋儗僼僩傪傕偭偰偄傞偑丄僋儗僼僩偺抂偑杽傑偭偰偄傞偨傔丄摐嵔偺枛抂晹暘偵偟偐寢崌偑偱偒側偄傕偺偱偁傞丅CBM9偑偦偺戙昞椺偱偁傞丅
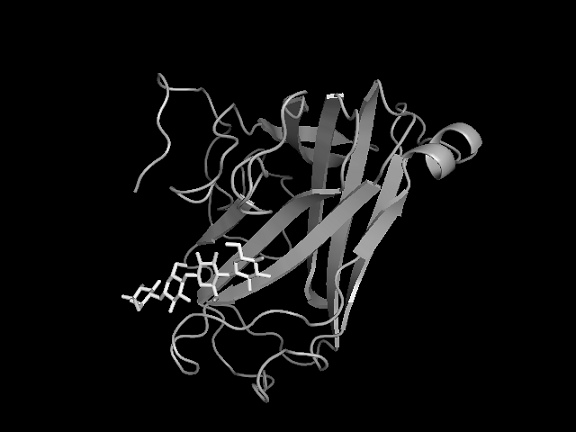
CBM偺僞儞僷僋幙偲偟偰偺峔憿偺傎偲傫偳偑丄β-僶儗儖峔憿偱丄偦偺懠偵丄儗僋僠儞偺傛偆側僩儗僼僅僀儖峔憿偑抦傜傟偰偄傞6,7)丅恾1偵CBM28偺峔憿傪帵偟偨丅β-僶儗儖峔憿傪桳偟偨僞僀僾B偺CBM偱偁傞8)丅
![]()
係丏CBM偺婡擻
峺慺偑CBM傪傕偮偙偲偺堄媊偲偟偰偼丄婎幙昞柺偵寢崌偡傞偙偲偵傛傝丄峺慺偺桳岠婎幙擹搙傪忋徃偝偣傞岠壥偑峫偊傜傟傞丅堦斒偵丄峺慺偼丄婎幙擹搙偺忋徃偵敽偄斀墳懍搙偑憹壛偡傞丅僙儖儔乕僛偺応崌丄婎幙偱偁傞僙儖儘乕僗偼悈偵晄梟惈偱偁傞偨傔偵丄婎幙昞柺偵寢崌偡傞偙偲偑丄峺慺偺婎幙擹搙傪忋偘傞偙偲偵側傞丅傑偨丄CBM傪傕偮峺慺偺懡條惈傪峫偊傞偲丄昁偢偟傕丄怗攠偺婎幙摿堎惈偲CBM偺昗揑偑摨偠偱偁傞昁梫偼側偔丄CBM偺昗揑偵寢崌偡傞偙偲偱丄杮棃偺婎幙偵傛傝嬤偯偗傞偲偄偆棙揰傪傕偭偰偄傞偲峫偊傜傟傞丅
5丏CBM偺寢崌摿惈偺夝愅
5-1丂CBM偼丄偳偙偵寢崌偡傞偺偐丠
僙儖儘乕僗旝寢徎慇堐偺儗儀儖偱丄僞僀僾A偵懏偡傞CBM1傗CBM3偼丄僙儖儘乕僗寢徎偺110柺偲屇偽傟傞慳悈柺偵寢崌偡傞9)丅偙傟偼丄崅懍尨巕娫椡尠旝嬀偱娤嶡偝傟偨T. reesei Cel7A偺摦偒偲傕堦抳偟偰偍傝丄CBM1傪傕偮峺慺偼丄寢徎惈僙儖儘乕僗偺110柺偵寢崌偟丄婎幙暘夝傪峴偭偰偄傞10)丅嵟嬤偺僐儞僺儏乕僞偵傛傞暘巕僔儈儏儗乕僔儑儞偱傕丄摨偠寢壥偑摼傜傟偰偄傞11)丅
堦曽丄Bae傜偼丄C. stercorarium偺CBM6偲R. albus Cel5B偺梈崌僞儞僷僋幙傪梡偄偰丄傾儖僼傽儖僼傽偺嵶朎暻傊偺寢崌傪嬥僐儘僀僪偱昗幆偟偨峈懱傪梡偄偰挷傋偨丅偦偺寢壥丄堐娗懇嵶朎偺嵶朎暻偵偼偁傑傝寢崌偣偢丄庡偵廮嵶朎偺嵶朎暻偵摿堎揑偵寢崌偡傞偙偲傪尒偄偩偟偨12)丅傑偨丄CBM偵His僞僌傪偮偗偰敪尰偝偣丄寀岝昗幆偟偨峈僸僗僞僌峈懱傪梡偄偨専弌傗13)丄CBM偲寀岝僞儞僷僋幙偲偺梈崌僞儞僷僋幙傪敪尰偡傞偙偲偱丄寀岝尠旝嬀壓偵偍偄偰丄CBM偺寢崌偡傞晹埵傪摿掕偡傞偙偲偑偱偒傞14)丅
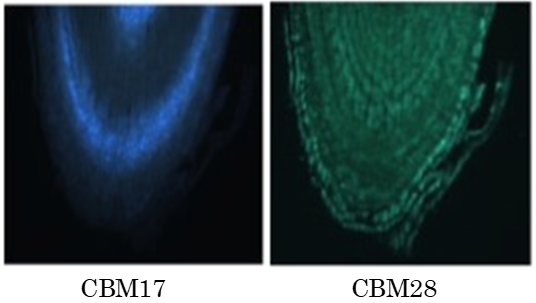
偙傟傜偺寢壥偐傜丄CBM偼丄怉暔嵶朎暻偵摿堎揑偵寢崌偡傞偑丄嵶朎暻偺応強傗丄嵶朎偺庬椶偵傛傝寢崌応強偑堎側偭偰偄傞偙偲偑柧傜偐偵側傝丄側偤CBM偑懡條側偺偐傪愢柧偟偰偄傞15)丅偨偲偊偽丄Clostridium josui偵桼棃偡傞CBM17偲CBM28偼丄僙儘僆儕僑摐偵懳偡傞寢崌摿惈偱偼丄帡偨惈幙傪帵偡傕偺偺丄寀岝僞儞僷僋幙偲偺梈崌僞儞僷僋幙傪梡偄偰丄怉暔嵶朎暻偺寢崌晹埵傪娤嶡偡傞偲柧傜偐偵堎側傞晹埵偵寢崌偟偰偄偨丅傑偨丄寢崌偟傗偡偄嵶朎暻偑嵶朎偺庬椶偵傛偭偰堎側偭偰偄偨 (恾2)16)丅
![]()
幚僶僀僆儅僗偼丄偦偺傑傑偺忬懺偱偼丄峺慺摐壔偡傞偙偲偑偱偒側偄偺偱丄慜張棟偑昁梫偵側傞丅偦偺慜張棟傪昡壙偡傞偨傔偵丄摐壔帋尡傪偟側偗傟偽側傜側偄偑丄CBM偺寢崌検偑懡偄帋椏偼丄摐壔棪傕崅偄偲偄偆憡娭偑偁傝丄摐壔帋尡偺慜偵暘夝偟傗偡偝傪昡壙偱偒傞17)丅傑偨丄僙儖儘乕僗寢徎壔搙偺曄壔傪FT-IR傗X慄夞愜傛傝傕姶搙傛偔丄CBM偺寢崌偼丄儌僯僞乕偡傞偙偲偑偱偒傞18)丅
5-2丂CBM偺寢崌偺僷儔儊乕僞
CBM偺摐幙傊偺寢崌偼丄偄偔偮偐偺僷儔儊乕僞偵傛偭偰昡壙偱偒傞丅偨偲偊偽丄寢崌掕悢傗嵟戝寢崌検偱偁傞丅婎杮揑偵偼丄寢崌夝愅偵偼Langmuir偺摍壏媧拝嬋慄偑巊梡偝傟偰偍傝丄偦偺媡懳悢僾儘僢僩偐傜丄寢崌掕悢Ka偲嵟戝寢崌検傪嶼弌偡傞偙偲偑偱偒傞19)丅偙傟偼丄堎側傞擹搙偺摐幙寢崌儌僕儏乕儖梟塼偵丄晄梟惈摐幙傪壛偊丄寢崌偟側偐偭偨忋惔偺枹寢崌晹暘偺僞儞僷僋幙擹搙傪掕検偡傞偙偲傛傝丄寢崌偟偨僞儞僷僋幙検傪嶼弌偟丄寢崌検傪悇掕偡傞傕偺偱偁傞丅寢崌掕悢偼丄偍偍傛偦丄105乣106µmol-1 偺僆乕僟乕偵偁傞丅嵟嬤偵側傝丄Sugimoto傜偼丄偙偺夝愅偵Hill偺幃傪摫擖偟偨丅廬棃偺Langmuir偺摍壏媧拝幃偱偺夞婣偱偼丄堎側傞寢崌掕悢傪傕偮寢崌晹埵偑暋悢懚嵼偟側偄偲夞婣偱偒側偐偭偨偑丄偙偺栤戣傪夝寛偟偨20)丅
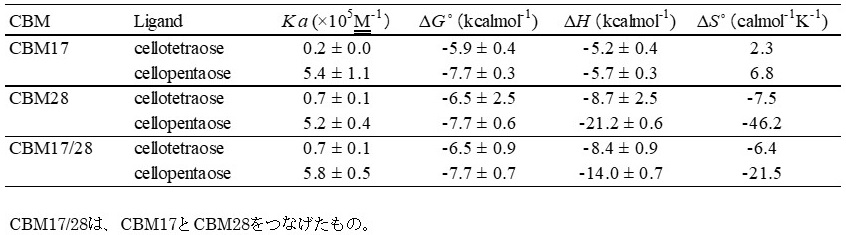
堦曽丄僞僀僾B偺傛偆側壜梟惈偺婎幙偵傕寢崌偱偒傞傛偆側CBM偺応崌丄摍壏揌掕擬検寁 (ITC) 偑昡壙偵巊梡偝傟傞丅偙傟偼丄CBM梟塼偵丄寢崌婎幙偱偁傞僆儕僑摐傪揌壓偡傞偙偲偱丄寢崌偺嵺偵敪惗偡傞傢偢偐側敪擬検傪懆偊應掕偡傞丅偙傟偵傛傝丄僞儞僷僋幙偲婎幙偲偺寢崌掕悢丄寢崌斾丄敪擬検偑摼傜傟丄僄儞僩儘僺乕曄壔傪嶼弌偡傞偙偲偑偱偒傞丅壜梟惈婎幙偺応崌偺寢崌掕悢偼丄104乣106偺僆乕僟乕偵偁傝丄婎幙偺挿偝偵傛偭偰曄壔偟丄捠忢挿偄傕偺傎偳丄崅偄寢崌掕悢傪帵偡丅昞2偵CBM17偲CBM28偺ITC偐傜摼傜傟偨寢崌僷儔儊乕僞傪帵偟偨丅CBM17偲28傪斾妑偡傞偲摨偠傛偆側寢崌掕悢傪傕偭偰偄傞偑丄寢崌偵偍偗傞敪擬検偑堎側偭偰偄傞丅偙傟偼丄CBM17偱偼丄僄儞僩儘僺乕偺婑梌偑戝偒偄偙偲傪帵偟偰偄傞丅傑偨CBM17偲28偑偮側偑偭偨傕偺偱偼丄僥僩儔僆乕僗偼丄2暘巕寢崌偱偒傞偑丄儁儞僞僆乕僗偼丄1暘巕偟偐寢崌偑偱偒側偄21)丅
昞2丂摍壏揌掕擬検寁偵傛傞CBM17偲28偺寢崌摿惈
俇丏CBM偺懴擬惈偺婑梌
偄偔偮偐偺僪儊僀儞傪奜偡偲怗攠偺懴擬惈偑掅壓偡傞偲偄偆曬崘偑偁傝丄偙偺傛偆側僪儊僀儞傪擬埨掕壔僪儊僀儞偲屇傫偱偄偨22)丅偙偺傛偆側僪儊僀儞偺傂偲偮偑CBM22偱偁傝丄屻偵CBM偱偁傞偙偲偑敾柧偟偨丅堦斒偵丄CBM偼β-僶儗儖峔憿傪偟偰偍傝丄怗攠僪儊僀儞傛傝傕僐儞僷僋僩偱丄擬埨掕惈偑崅偄丅C. stercorarium Xyn10B偺応崌丄帵嵎憱嵏擬検寁偱曄惈壏搙傪應掕偡傞偲丄GH10偺怗攠僪儊僀儞 (CM10) 偺曄惈壏搙偼74亷偱偁偭偨丅堦曽CBM22偺曄惈壏搙偼82亷偵偁傝丄柧傜偐偵怗攠傛傝傕擬偵埨掕側峔憿偱偁偭偨丅偙偺CBM傪梈崌偟偨峺慺CBM22-CM10偼丄棟桼偼柧傜偐偱偼側偄偑丄曄惈壏搙偑6亷忋徃偟丄80亷偵側傝丄壏搙偵懳偟偰僞儞僷僋幙慡懱偺峔憿埨掕壔傪堷偒婲偙偟偨丅偙傟偼丄怗攠僪儊僀儞偲CBM偲偺娫偺憡屳嶌梡偵傛傝丄僞儞僷僋幙慡懱偑擬偵懳偟偰埨掕壔偟偨偲峫偊傜傟傞 (恾3)23)丅
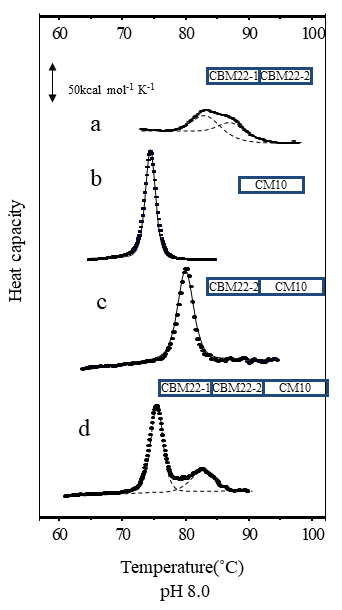
恾3丂CBM傪傕偮峺慺偺曄惈壏搙
俈丏CBM偺婎幙曵夡嶌梡
CBM偼丄慇堐昞柺偵寢崌偡傞偩偗偱側偔丄昞柺偺峔憿傪旕壛悈暘夝揑偵曵夡偝偣偰偄傞丅僙儖儘乕僗偺応崌丄僙儘僿僉僒僆乕僗傑偱偼丄悈偵壜梟偱偁傞偑丄偦傟埲忋偺挿嵔偺応崌偼悈偵梟偗側偄丅偟偐偟丄峺慺偑僙儖儘乕僗傪暘夝偡傞偵偼丄彮側偔偲傕僋儗僼僩撪晹傑偱丄梀棧偟偨僙儖儘乕僗嵔偑撏偔昁梫偑偁傞丅偦偙偱丄摐幙寢崌儌僕儏乕儖偼丄寢徎柺偐傜偺僙儖儘乕僗嵔偺攳棧偵岠壥偑偁傞偲峫偊傜傟偰偒偨丅幚嵺偵丄摐幙寢崌儌僕儏乕儖偲柸慇堐傪僀儞僉儏儀乕僔儑儞偡傞偲丄柸慇堐昞柺偵偝偝偔傟偺傛偆側曵夡偑尒傜傟傞24)丅偝傜偵丄僙儖儘乕僗慇堐偺暘巕撪悈慺寢崌丄嵔娫偺悈慺寢崌偑摐幙寢崌儌僕儏乕儖偺寢崌偵傛傝庛傔傜傟偰偍傝25)丄偙偺嶌梡傪僶僀僆儅僗偺慜張棟偵壛偊傞偙偲偵傛傝摐壔棪傪忋偘傞偙偲偑偱偒偰偄傞26)丅
俉丏CBM傛傞怉暔昦奞偺墳摎
CBM偼丄旝惗暔偩偗偱側偔怉暔偵傕暘晍偟偰偍傝丄慜弎偺CBM49偺懠丄CBM43偼壴暡娗僞儞僷僋幙偵尒偮偐偭偰偄傞27)丅怉暔昦尨嬠偵偼丄僙儖儘乕僗偵寢崌偡傞CBM1傪僞儞僨儉偵偮側偄偩僞儞僷僋幙傪惗嶻偡傞傕偺偑偁傝丄偙傟傜偺CBM1偼丄怉暔偵懳偟偰杊屼墳摎傪堷偒偙偡偙偲偑抦傜傟偰偄傞28)丅偦偙偱丄CBM3偺梟塼傪僩儅僩偵暚柖偟丄僩儅僩惵屚傟昦嬠傪娷傫偩攟搚偵堏怉偟偨偲偙傠丄CBM3梟塼傪暚柖偟偨僩儅僩偼屚傟偢丄昦尨嬠偵懳偟偰掞峈惈傪帵偟偨 (恾4)29)丅偙偺傛偆偵丄CBM梟塼傪暚柖偟丄CBM偑怉暔偵寢崌偡傞偙偲偵傛傝丄怉暔懁偵壗傜偐偺僔僌僫儖偑揱傢傝丄偦傟偵傛傝怉暔偑杮棃傕偭偰偄傞昦尨旝惗暔偵懳偡傞掞峈惈傪桿摫偡傞偙偲偑傢偐偭偨30)丅


俋丏偍傢傝偵
CBM偼丄扨偵丄怗攠傪婎幙偵嬤偯偗傞偲偄偆岠壥傪傕偮偩偗偱側偔丄條乆側婡擻傪傕偮偙偲偑暘偐偭偰偒偨丅CBM偑偮偔偙偲偱丄峺慺偺妶惈傗婎幙摿堎惈偑塭嬁傪庴偗傞偙偲丄CBM帺恎偑寢徎惈僙儖儘乕僗偵懳偟偰曵夡妶惈傪傕偮偙偲傕柧傜偐偵側偭偰偒偨丅傑偨丄僨乕僞儀乕僗偵偼懡偔偺峔憿偺堎側傞CBM偑懚嵼偟偰偍傝丄側偤偙偺傛偆側懡條側CBM偑懚嵼偟偰偄傞偐傪峫偊傞偲丄変乆偑抦傜側偄傛偆側嵶朎暻偺峔憿偺嵎堎偵旝惗暔偼懳墳偡傞偨傔偵丄CBM傪恑壔偝偣偨偲傕峫偊傞偙偲偑偱偒傞丅旝惗暔僎僲儉偺夝愅偑恑傓偙偲偵傛傝丄崱屻傕怴偨側CBM偑敪尒偝傟傞偲梊憐偝傟傞丅堦曽丄CBM偺墳梡椺偺傂偲偮偲偟偰徯夘偟偨偑丄惗偒偨怉暔偺嵶朎暻偵CBM偑寢崌偡傞偙偲偱丄怉暔懁偵杊屼墳摎傪桿摫偱偒傞偙偲偑傢偐偭偨丅偙偺懠偵傕丄僞僌偲偟偰偺棙梡傗丄CBM傪敪尰偟偨旝惗暔偺僙儖儘乕僗慇堐傊偺屌掕壔丄CBM偲偺楢寢偵傛傝摿掕偺暔幙傪僙儖儘乕僗忋偵屌掕偡傞偲偄偭偨墳梡椺傕偁傞31)丅峺慺杮懱偺怗攠妶惈偼傕偪傠傫廳梫偱偁傞偑丄偙偆偟偨晅壛僪儊僀儞偺傕偮栶妱傕堄枴偑偁傞偙偲傪帺慠偼嫵偊偰偄傞丅
幱帿
杮尋媶偺堦晹偼丄壢妛尋媶旓曗彆嬥20580362丄埳摗婰擮嵿抍丄嶰廳戝妛妛撪COE-B偺彆惉偵傛傝峴傢傟傑偟偨丅
暥專
1) VanTibeurgh, H., Tomme, P., Claeyssens, M., Bhikhabhai, R., Pettersson, G.: FEBS Lett., 204, 223 (1986).
2) Gilkes, N. R., Henrissat, B., Kilburn, D. G., Miller Jr, R. C., Warren, R. A. J.: Microbiol. Rev., 55, 303 (1991).
3) Cantarel, B. L., Coutinho, P. M., Rancurel, C., Berrnard, T., Lombard, V., Henrissat, B.: Nucleic Acids Res., 37, D233 (2009).
4) Herve, C., Rogowski, A., Blake, A. W., Marcus, S. E., Gilbert, H. J., Knox, J. P.: Proc. Natl. Acad. Sci. USA, 107, 15293 (2010).
5) Boraston, A. B., Bolam, D. N., Gilbert, H. J., Davies, G. J.: Biochem. J., 382, 769 (2004).
6) Hashimoto, H.: Cell. Mol. Life Sci., 63, 2954 (2006).
7) Guillen, D., Sanchez, S., Rodriguez-Sanoja, R.: Appl. Microbiol. Biotechnol., 85, 1241 (2010).
8) Tsukimoto, K., Takada, R., Araki, Y., Suzuki, K., Karita, S., Wakagi, T., Shoun, H., Watanabe, T., Fushinobu, S.: FEBS Lett., 584, 1205 (2010).
9) Lehtio, J., Sugiyama, J., Gustavssion, M., Fransson, L., Linder, M., Teeri, T. T.: Proc. Natl. Acad. Sci. USA, 100, 484 (2003).
10) Igarashi, K., Uchihashi, T., Koivula, A., Wada, M., Kimura, S., Okamoto, T., Penttila, M., Ando, T., Samejima, M.: Science, 333, 1279 (2011).
11) Nimlos, M. R., Beckham, G. T., Mtthews, J. F., Bu, L., Himmel , M. E., Crowley, M. F.; J. Biol. Chem., 287, 20603 (2012).
12) Bae, H.-J., Turcott, G., Chamberland, H., Karita, S., Vezina, L.-P.: FEMS Microbiol. Lett., 227, 175 (2003).
13) Knox, J. P.: Curr. Opin. Plant Biol., 11, 308 (2008).
14) Ding, S.-Y., Xu, Q., Ali, M. K., Baker, J. O., Bayer, E. A., Barak, Y., Lamed, R., Sugiyama, J., Rumbles, G., Himmel, M. E.: BioTechniques, 41, 435 (2006).
15) Blake, A. W., McCartney, L., Flint, J. E., Bolam, D. N., Boraston, A. B., Gilbert, H. J., Knox, J. P.: J. Biol. Chem., 281, 29321 (2006).
16) Araki, Y., Karita, S., Tsuchiya, T., Kondo, M., Goto, M.: Biosci. Biotechnol. Biochem., 74, 802 (2010).
17) Kawakubo, K., Karita, S., Araki, Y., Watanabe, S., Oydomari, M., Takada, R., Tanaka, F., Abe, K., Watanabe, T., Honda, Y., Watanabe, T.: Biotechnol. Bioeng., 105, 499 (2009).
18) Kljun, A., Benians, T. A., Goubet, F., Meulewaeter, F., Knox, J. P., Blackburn, R. S.: Biomacromol., 12, 4121 (2011).
19) Gilkes, N. R., Jervis, E., Henrissat, B., Tekant, B., Miller Jr, R. C., Warren, R. A. J., Kilburn, D. G.: J. Biol. Chem., 267, 6743 (1992).
20) Sugimoto, N., Igarashi, K., Wada, M., Samejima, M.: Langmuir, 28, 14323 (2012).
21) Araki, Y., Karita, S., Tanaka, A., Kondo, M., Goto, M.: Biosci. Biotechnol. Biochem., 73, 1028 (2009).
22) Fontes, C. M., Hazlewood, G. P., Morag, E., Hall, J. B., Hirst, H., Gilbert, H. J.: Biochem. J., 307, 151 (1995).
23) Araki, R., Karita, S., Tanaka, A., Suzuki, M., Kimura, T., Sakka, K.: Biosci. Biotechnol. Biochem., 72, 1322 (2007).
24) Din, N., Gilkes, N. R., Tekant, B., Miller Jr, R. C., Warren, R. A. J., Kilburn, D. G.: Bio/Technol., 9, 1096 (1991).
25) Hall, M., Bansal, P., Lee, J. H., Realff, M. J., Bommarius, A. S.: Bioresource Technol., 102, 2910 (2011).
26) Ciolacu, D., Kovac, J., Kokol, V.: Carbohydr. Res., 345, 621 (2010).
27) Barral, P., Suarez, C., Batanero, E., Alfonso, C., Alche, J. D., Rodriguez-Garcia, M. I., Villalba, M., Rivas, G., Rodriguez, R.: Biochem. J., 390, 77 (2006).
28) Gaulin, E., Drame, N., Lafitte, C., Torto-Alalibo, T., Martinez, Y., Ameline-Torregrosa, C., Khatib, M., Mazarguil, H., Villalba-Mateos, F., Kamoun, S., Mazars, C., Dumas, B., Bottin, A., Esquerre-Tugaye, M.-T., Rickauer, M.: Plant Cell, 18, 1766 (2006).
29) 姟揷廋堦丄惔悈彨暥丗怉暔杊塽, 66, 326 (2012).
30) Dumas, B., Bottin, A., Gaulin, E., Esquerre-Tugaye, M.-T.: Trends Plant Sci., 13, 160 (2008).
31) Shoseyov, O., Shani, Z., Levy, I.: Microbiol. Mol. Biol. Rev., 70, 283 (2006).
![]() 丂
丂