【トピックス】
ホタルの進化は茶道のこころ
大場裕一
名大院・生命農学
茶の湯の秘伝書・山上宗二記 (やまのうえのそうじき) には「習うは古を専らに用うべし、作意は新しきを専らとす」とある1)。千利休 (1522~1591) は、京の桂川で漁夫が使っていた魚籠 (びく) を花入 (はないれ) に見立て、茶室に用いたという (図1)。このように、古くからあるものの価値を踏襲しながら用途の転換を試み、そうすることで思いがけない新しい美を生み出す「見立て」の精神は、古来からの日本の美意識であった。ところが、この創造美の実践者は桃山の茶人たちが最初ではなかった。

図1 桂籠 (高さ19cm 筆者個人蔵)
夏の夜の川辺に舞うホタル (図2) の話である。ホタルの発光は、古くから歌に詠まれ、また科学者の興味の対象でもあった。半世紀におよぶ詳細な発光メカニズム研究の結果、その反応は図3に示すようなルシフェラーゼ (酵素) によるルシフェリン (基質) の酸化であることが明らかとなっている。光は、酸化反応により生成した励起状態のオキシルシフェリンが基底状態に戻るさいに放出される。ところで、このような巧妙なシステムは一体どのような道筋で進化したのだろうか。本編では、“ホタルのルシフェラーゼの進化には、思いがけない自然の「見立て」が関与していたかもしれない”という我々の最近の研究から得られた仮説を紹介したい (進化という視点が、酵素の本質を知るなんらかの手がかりを与えるであろうことを期待しつつ)。

図2 ゲンジボタルLuciola cruciata (体長約14mm 岐阜県美濃市)
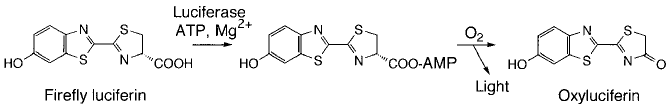
図3 ホタルの発光反応メカニズム
酵素工学研究会の会員諸氏にはあらためて申すまでもないが、酵素の触媒作用は実に多種多様で興味が尽きない。しかし、その中でも「光を出す酵素」ほど奇妙なモノもないのではなかろうか。そういう思いから、私はルシフェラーゼの進化に興味を持ち、これまで研究を行なってきた。結論を先に述べてしまうと、我々は最近“ホタルルシフェラーゼは脂肪酸代謝に関わる酵素のひとつから進化した”ことの確からしい証拠を見つけた。おそらくどんな生物でも持っているだろう脂肪酸代謝酵素から、ルシフェラーゼという特殊な酵素への転換─それが本当ならば、これは実に大胆な自然の「見立て」と言うべきではないか。
光る生き物は、もちろんホタルだけではない。他によく知られているものに、ウミホタルや夜光虫、ホタルイカなどがあるが、これらの発光もホタルと同様に酵素反応 (ルシフェリン-ルシフェラーゼ反応) であることがわかっている。しかし、(あまり知られていないことだが) これら発光生物のルシフェリンの化学構造 (図4) やルシフェラーゼのアミノ酸配列には、お互い類似性が見られない。このことから、発光とは生物の進化過程で独立に何度も獲得されてきた形質であると考えられており、ホタルもその例外ではない。つまり、ホタルのルシフェラーゼは昆虫の中で独自に進化した酵素であり、ウミホタルなどのルシフェラーゼとは起源的になんの関係もないのである。
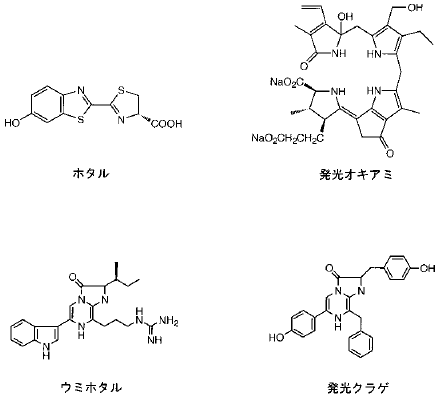
図4 さまざまな発光生物のルシフェリン
ホタルルシフェラーゼの起源に関係がありそうなのは、むしろ発光とはまったく縁のなさそうな酵素である。これまで知られているものの中で最もアミノ酸配列が似ているのはクマル酸CoAリガーゼという酵素で2)、植物のみに知られるこの酵素は、樹木の木質を作るリグニンの生合成などに関与している。また、クマル酸CoAリガーゼほどではないが、ルシフェラーゼは脂肪酸CoA合成酵素とも高いアミノ酸相同性を示す。この酵素は、β酸化における最初のステップを触媒する酵素であり、その存在はバクテリアから哺乳類まで知られている。このように機能に着目すると、この2つの酵素とルシフェラーゼには何のつながりもなさそうに思われる。しかし、触媒反応の詳細に着目するとその関連性が見えてくる (図5)。これらの酵素はすべてATPとマグネシウム要求性であり、しかも反応の最初のステップが共通 (カルボン酸のアデニル化) なのである (注1)。このようにアミノ酸配列および触媒様式から推定する限り、ルシフェラーゼは何らかの「カルボン酸のアデニル化酵素」から進化した可能性が想像される。しかし、それ以上のことを議論するにはデータが不足であった。
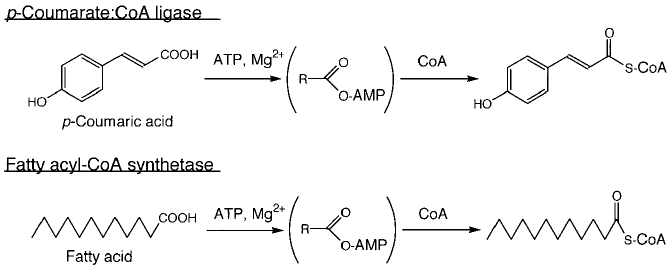
図5 クマル酸CoAリガーゼと脂肪酸CoA合成酵素の触媒反応
ある生物の持つ形質について進化を推定するさいに最も有効な方法は、その生物自身の中に過去の痕跡を見つけ出すことである。たとえば、ニシキヘビに見られる痕跡的な腰帯やウマの四肢にある小さな第二・第四中手骨などは、その生物がたどってきた道筋を示す格好の証拠である (注2)。しかし、このような進化の痕跡がいつも都合良く見つかるとは限らない。そうした場合には、まず近縁な生き物を調べて比較してみるのが常法である。ホタルの進化を考えるために、我々は非発光性の昆虫からルシフェラーゼに似た遺伝子を見つけ、その遺伝子機能を解析する研究計画を立てた。しかしその前に、もしかするとホタルルシフェラーゼ自体に痕跡的な過去の触媒活性が残っているかもしれないと思い、調べてみることにした。実験の結果、ルシフェラーゼがオレイン酸をCoAエステル化する活性を持つことがわかった3)。さらに、32PATPを用いて基質選択性を調べたところ、脂肪酸はリノレン酸やアラキドン酸などの長鎖で不飽和度の高いものほど基質に適しており、炭素数14以下の中・短鎖のものは基質にならないことがわかった。また、クマル酸CoAリガーゼの基質であるクマル酸やカフェイン酸に対する活性もなかった (図6)。「ルシフェラーゼは発光活性に特化した特別な酵素である」と勝手に想像していた私にとって、この結果は驚きであった。では、この脂肪酸CoA合成酵素活性がルシフェラーゼの過去の触媒活性の名残りと考えてよいのだろうか。
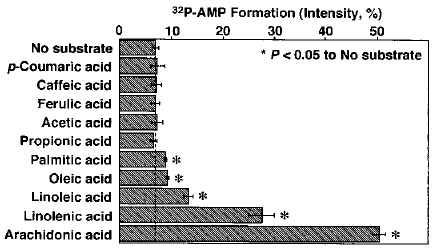
図6 ルシフェラーゼの基質選択性
(数値はルシフェリンを100としたときの値)
そこで次に、ホタルに近縁だが光らない昆虫としてキイロショウジョウバエ (Drosophila melanogaster) を調べてみることにした。むろんホタルにもっと近縁な昆虫はたくさんあるが、研究しやすさではショウジョウバエには及ばない。さっそくショウジョウバエの全ゲノムデータをサーチしてみたところ4)、ホタルルシフェラーゼに非常によく似た遺伝子がひとつだけあることがわかった。この遺伝子は機能が未知で、単に“CG6178”というIDが付けられていた。分子系統解析を行なってみると、このCG6178が確かにホタルルシフェラーゼのオルソローグ (共通祖先の種分化に由来する遺伝子) であることがわかった (図7)。つまりCG6178 は、これまで知られている遺伝子のなかで最もホタルルシフェラーゼに近い「ルシフェラーゼではない」遺伝子なのである。しかもホタルと同じ昆虫類の遺伝子なので、その機能を明らかにすることは進化を考える上で非常に意義がある。さっそくCG6178遺伝子をクローニングし、大腸菌に発現させた。タンパクを精製したのち、先と同様の酵素活性試験を行なってみたところ、(期待どおり!) 脂肪酸に対するCoAエステル化の活性が確認された。ちなみに、CG6178タンパクをルシフェリンと混合してみたが、発光活性は見られなかった。また、ルシフェリンをCoAエステル化する活性もなかった。これにより、ルシフェラーゼ様遺伝子CG6178はショウジョウバエの脂肪酸CoA合成酵素であると結論づけられた5)。
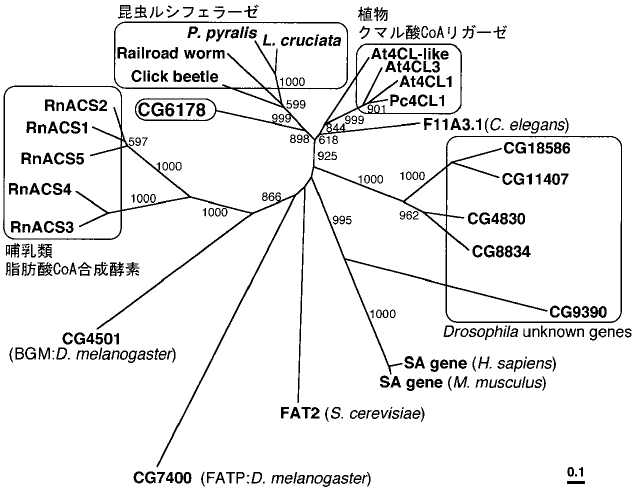
図7 アミノ酸配列に基づく分子系統解析 (ref. 5 より一部改変)
以上の結果は、ホタルルシフェラーゼが脂肪酸CoA合成酵素から進化したことを強く示唆するものである。しかし、なぜルシフェラーゼは祖先の機能である脂肪酸CoA合成酵素活性を保持しつづけたのだろう。もしかすると、現在もホタルはルシフェラーゼを脂肪酸CoA合成酵素として使用しているのかもしれない。
ホタルの進化について、ひとつまだ触れていなかった問題が「ルシフェリンの由来」についてである。実は、ホタルのルシフェリンがどのようにして生合成されるのか、また、もともと何に使われていた分子だったのかなどは一切わかっていない。しかし私は、ホタルの進化を知る上でルシフェリンの由来を知ることは、もしかするとルシフェラーゼの起源よりもずっと重要な問題なのではないかとひそかに思っている。というのは実は、ホタルルシフェラーゼは条件しだいで (例えば、DMSO中でt-BuOKを加えると) ルシフェラーゼがなくても発光するのである (ただし、発光効率は格段に落ちるが)。つまりルシフェリンとは、それ自体「発光に適した物質」だったのだ。
発光への道は、ホタルの祖先がルシフェリンを生合成するようになったとき (あるいは食物から調達してもいいのだが) 初めて開けたに違いない。それからどういうわけか、発光とは縁もなかった脂肪酸CoA合成酵素がルシフェリンの発光性を触媒する酵素としてリクルートされ、その後、変異と淘汰が繰り返された結果、現在のルシフェラーゼが完成した。
しかし、ホタルの祖先はなぜ脂肪酸CoA合成酵素をルシフェラーゼに選んだのだろうか?発光という酸化反応を触媒するのなら、オキシダーゼの方がいいように思えるのだが・・・。まあ、おそらくそのあたりが私のセンスの限界のようだ。自然はいつも我々の予想も及ばないような大胆さでさまざまな用途の転換を繰り返し、常に目映いばかりの多様性を生み出し続けている。
利休のすぐれた「見立て」のセンスにより、その桂籠は花入の定番として今に伝えられている。さて、世界に2000種以上とも言われるホタル科の成功も、いにしえの酵素の「見立て」がきっかけだったのだろうか? この進化の秘伝書は、まだひもとかれたばかりである。
注
注1:しかし、アデニル化よりあとのステップは、これらの酵素で大きく異なっている。ルシフェラーゼによる発光反応では、アデニル化に続いて酸素分子の付加が起こり“ジオキセタノン”とよばれる不安定な中間体を経て、オキシルシフェリンができる。一方、クマル酸CoA リガーゼと脂肪酸CoA合成酵素では、カルボン酸に付いていたAMPがコエンザイムAと置き換わり、CoAエステルができる。
注2:これらのちっぽけな骨のカケラは、ヘビの祖先に後肢があったこと、ウマの祖先に示指と薬指があったこと、を雄弁に語っている。
文献
1) 「山上宗二記」日本の茶書1 (東洋文庫、平凡社、1971) 茶湯者覚悟十体より抜粋
2) Schröder, J.: Nucleic Acids Res., 17, 460 (1989).
3) Oba, Y., Ojika, M., Inouye, S.: FEBS Lett., 540, 251 (2003).
4) Fly Base (http://flybase.net/)
5) Oba, Y., Ojika, M., Inouye, S.: Gene, 329, 137 (2004).
![]()